Aminoacyl-ARNt synthétase

Glutaminyl-ARNt synthétase d'E. coli complexée avec un ARNtGln (PDB 1QRS[1]).
Les aminoacyl-ARNt synthétases (abréviation aaRS) forment une famille de ligases (EC ) qui catalysent l'estérification des acides aminés protéinogènes (abréviation aa) sur l'extrémité 3' des ARN de transfert (abréviation ARNt). Cette étape est essentielle à la traduction des ARN messagers en protéines, les acides aminés ainsi ajoutés à l'extrémité des ARNt étant incorporés par les ribosomes dans la chaîne polypeptidique en cours de synthèse. Ce sont des protéines essentielles, conservées chez tous les êtres vivants.
Chez la plupart des organismes, il existe une aminoacyl-ARNt synthétase pour chacun des acides aminés protéinogènes, hormis pour la sélénocystéine, liée à son ARN de transfert par un mécanisme indirect passant par la sérine. Chacune de ces enzymes reconnaît un acide aminé et un ou plusieurs ARNt isoaccepteurs. Leur fonction est essentielle à la fidélité de la traduction du code génétique, car c'est elles qui garantissent que l'acide aminé qui est ainsi estérifié à l'extrémité de l'ARNt correspond bien au bon anticodon. Aucune vérification n'est ensuite effectuée au niveau du ribosome.
Sommaire
1 Mécanisme
2 Classes d'aminoacyl-ARNt synthétases
3 Structures
4 Relecture
5 Notes et références
6 Voir aussi
6.1 Articles connexes
Mécanisme |
Les aminoacyl-ARNt synthétases catalysent la réaction d'aminoacylation en deux étapes[2], elles activent d'abord l'acide aminé en formant un aminoacyl-adénylate avec l'ATP, avec libération de pyrophosphate. L'acide aminé ainsi activé reste lié à l'enzyme et est transféré sur la fonction 2'-OH ou 3'-OH du ribose de l'adénosine située à l'extrémité 3' de l'ARNt :
acide aminé + ATP → aminoacyl-AMP + pyrophosphate ;
aminoacyl-AMP + ARNt → aminoacyl-ARNt + AMP.
La plupart des aminoacyl-ARNt synthétases peuvent réaliser la première de ces deux réactions en l'absence de l'ARNt, mais la glutamyl-ARNt synthétase, la glutaminyl-ARNt synthétase, l'arginyl-ARNt synthétase et la lysyl-ARNt synthétase I, en sont incapables si l'ARNt est absent. On dit que l'activation de ces acides aminés (glutamate, glutamine, lysine, arginine) est ARNt-dépendante.
Sur le plan de la nomenclature des enzymes, les aminoacyl-ARNt synthétases sont des ligases et sont classées EC . Elles se déclinent suivant la nature de l'acide aminé dont elles sont spécifiques : alanyl-ARNt synthétase pour l'alanine, méthionyl-ARNt synthétase pour la méthionine, etc.
Chez la bactérie Escherichia coli et la levure Saccharomyces cerevisiae (un eucaryote), il existe bien 20 aminoacyl-ARNt synthétases, une pour chaque acide aminé standard[3].
| Acide aminé | Enzyme | Abréviation | N° EC | Classe | ARNt substrat | ||
|---|---|---|---|---|---|---|---|
| Alanine | A | Ala | Alanyl-ARNt synthétase | AlaRS | EC | II | ARNtAla |
| Arginine | R | Arg | Arginyl-ARNt synthétase | ArgRS | EC | I | ARNtArg |
| Asparagine | N | Asn | Asparaginyl-ARNt synthétase | AsnRS | EC | II | ARNtAsn |
| Aspartate | D | Asp | Aspartyl-ARNt synthétase | AspRS | EC | II | ARNtAsp |
| Cystéine | C | Cys | Cystéinyl-ARNt synthétase | CysRS | EC | I | ARNtCys |
| Glutamate | E | Glu | Glutamyl-ARNt synthétase | GluRS | EC | I | ARNtGlu |
| Glutamine | Q | Gln | Glutaminyl-ARNt synthétase | GlnRS | EC | I | ARNtGln |
| Glycine | G | Gly | Glycyl-ARNt synthétase | GlyRS | EC | II | ARNtGly |
| Histidine | H | His | Histidyl-ARNt synthétase | HisRS | EC | II | ARNtHis |
| Isoleucine | I | Ile | Isoleucyl-ARNt synthétase | IleRS | EC | I | ARNtIle |
| Leucine | L | Leu | Leucyl-ARNt synthétase | LeuRS | EC | I | ARNtLeu |
| Lysine | K | Lys | Lysyl-ARNt synthétase | LysRS | EC | I / II | ARNtLys |
| Méthionine | M | Met | Méthionyl-ARNt synthétase | MetRS | EC | I | ARNtMeti (initiateur), ARNtMete (élongateur) |
| Phénylalanine | F | Phe | Phénylalanyl-ARNt synthétase | PheRS | EC | II | ARNtPhe |
O-Phosphosérine | - | Sep | O-Phosphoséryl-ARNt synthétase | SepRS | EC | II | ARNtCys |
| Proline | P | Pro | Prolyl-ARNt synthétase | ProRS | EC | II | ARNtPro |
| Pyrrolysine | O | Pyl | Pyrrolysyl-ARNt synthétase | PylRS | EC | II | ARNtPyl |
| Sérine | S | Ser | Séryl-ARNt synthétase | SerRS | EC | II | ARNtSer, ARNtSec |
| Thréonine | T | Thr | Thréonyl-ARNt synthétase | ThrRS | EC | II | ARNtThr |
| Tryptophane | W | Trp | Tryptophanyl-ARNt synthétase | TrpRS | EC | I | ARNtTrp |
| Tyrosine | Y | Tyr | Tyrosyl-ARNt synthétase | TyrRS | EC | I | ARNtTyr |
| Valine | V | Val | Valyl-ARNt synthétase | ValRS | EC | I | ARNtVal |
Bien que ces organismes modèles possèdent toutes les aminoacyl-ARNt synthétases, ce n'est pas le cas de tous les organismes. En effet, il n'est pas rare que la glutaminyl-ARNt synthétase (GlnRS) soit absente, notamment chez 90 % des bactéries, et chez toutes les archées. Il a par ailleurs été mis en évidence le fait que les mitochondries sont également dépourvues de cette enzyme. Chez la bactéries et les archées, l'asparaginyl-ARNt synthétase peut également être absente. Dans tous les cas (et comme la synthèse des deux aminoacyl-ARNt, l'Asn-ARNt(Asn) et le Gln-ARNt(Gln), est essentielle), les organismes utilisent une voie alternative, qui implique deux étapes enzymatiques successives[4].
Classes d'aminoacyl-ARNt synthétases |
Il existe deux classes d'aminoacyl-ARNt synthétases (aaRS), appelées classe I et classe II, de propriétés structurales et fonctionnelles très distinctes. Les aminoacyl-ARNt synthétases de classe I estérifient l'acide aminé sur le 2'-OH de l'adénosine terminale de l'ARNt, et se lient à ce dernier par le petit sillon du bras accepteur. Leur structure est organisée autour d'un domaine constitué d'un feuillet bêta parallèle, entouré d'hélices alpha appelé pli Rossmann et commun à de nombreuses enzymes utilisant un cofacteur nucléotidique (ATP, NAD+). Les aminoacyl-ARNt synthétases de classe II estérifient l'acide aminé sur le 3'-OH de l'adénosine terminale de l'ARNt et se lient par le grand sillon de son bras accepteur. Elles possèdent une structure construite autour d'un feuillet bêta antiparallèle [5],[6],[7].
À l'exception de la lysyl-ARNt synthétase, pour laquelle il existe des formes de classe I ou de classe II, suivant les espèces, pour tous les autres acides aminés, on ne trouve systématiquement qu'une forme d'enzyme, soit de classe I, soit de classe II. Ces deux classes d'enzymes structurellement distinctes semblent être le résultat d'un processus d'évolution convergente.
Parmi les enzymes de classe I, on trouve la leucyl-ARNt synthétase (LeuRS), l'isoleucyl-ARNt synthétase (IleRS), la valyl-ARNt synthétase (ValRS), la cystéinyl-ARNt synthétase (CysRS), la méthionyl-ARNt synthétase (MetRS), l'arginyl-ARNt synthétase (ArgRS), la glutamyl-ARNt synthétase (GluRS), la glutaminyl-ARNt synthétase (GlnRS), la lysyl-ARNt synthétase de type I (LysRSI), la tyrosyl-ARNt synthétase (TyrRS) et la tryptophanyl-ARNt synthétase (TrpRS).
Les enzymes de classe II comprennent les séryl-ARNt synthétase (SerRS), thréonyl-ARNt synthétase (ThrRS), histidyl-ARNt synthétase (HisRS), prolyl-ARNt synthétase (ProRS), glycyl-ARNt synthétase (GlyRS), alanyl-ARNt synthétase (AlaRS), aspartyl-ARNt synthétase (AspRS), asparaginyl-ARNt synthétase (AsnRS), lysyl-ARNt synthétase de type II (LysRSII), phénylalanyl-ARNt synthétase (PheRS), pyrrolysyl-ARNt synthétase (PylRS) et O-phosphoséryl-ARNt synthétase (SepRS).
Structures |
Les deux classes d'aminoacyl-ARNt synthétases sont des protéines multidomaines. Elles sont généralement constituées d'un domaine catalytique et d'un domaine de liaison à l'anticodon. Certaines d'entre elles possèdent également un autre domaine de liaison à l'ARN qui assure une fonction de relecture en clivant la liaison avec les acides aminés qui ne correspondent pas à l'ARN de transfert.
Le domaine catalytique de toutes les aminoacyl-ARNt synthétases sont homologues les uns avec les autres alors que les enzymes de classe I et de classe II ne sont pas apparentées. Celles de classe I possèdent le pli Rossmann ainsi qu'une architecture à brins β parallèles tandis que celles de classe II présentent un repliement particulier avec des brins β antiparallèles.
Le domaine en hélices α de liaison à l'anticodon des arginyl-, glycyl- et cystéinyl-ARNt synthétases est appelé domaine DALR en raison des acides aminés conservés (Asp, Ala, Leu et Arg) qui lui sont caractéristiques[8].
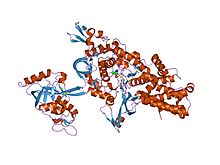
| Pfam | PF08264 |
|---|---|
| InterPro | IPR013155 |
| SCOP | 1ivs |
| SUPERFAMILY | 1ivs |
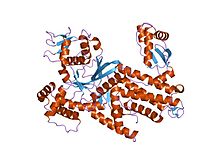
| Pfam | PF05746 |
|---|---|
| Clan Pfam | CL0258 |
| InterPro | IPR008909 |
| SCOP | 1bs2 |
| SUPERFAMILY | 1bs2 |

| Pfam | PF09190 |
|---|---|
| Clan Pfam | CL0258 |
| InterPro | IPR015273 |
Relecture |
Certaines aminoacyl-ARNt synthétases ont à discriminer des acides aminés structurellement très proches, comme l'isoleucyl-ARNt synthétase qui doit distinguer l'isoleucine de la valine, lesquelles ne diffèrent que par un groupe méthyle –CH3. La différence d'affinité entre le substrat naturel, l'isoleucine, et un substrat incorrect, en l'occurrence la valine, est insuffisante pour éviter que, parfois, la valine ne soit estérifiée sur l'ARNtIle par cette enzyme. Si cet évènement se produisait trop fréquemment, cela conduirait à l'incorporation fréquente de valine à la place d'isoleucine au niveau du ribosome, et donc à la synthèse de protéines anormales.
Ces aminocyl-ARNt-synthétases possèdent donc en général un mécanisme de relecture (en anglais : proofreading), qui permet à l'enzyme de vérifier, après la réaction, que l'acide aminé estérifié à l'extrémité de l'ARNt est bien l'acide aminé correct. Il existe sur l'enzyme un deuxième site de contrôle de l'acide aminé estérifié à l'ARNt. Si une erreur a été commise, la liaison ester formée est hydrolysée, ce qui permet d'effectuer une correction[9]. Cette correction d'un aminoacyl-ARNt incorrect correspond à un mécanisme dit post transfert, tout simplement parce que l'acide aminé incorrect a été transféré sur l'ARNt. Il existe également un autre type de correction, appelé "pré-transfert", et qui intervient lorsque l'acide aminé incorrect est activé en aminoacyl-adénylate. Ce dernier est alors hydrolysé avant son transfert sur l'ARNt[10].
Ce sont des mécanismes enzymatiques qui permettent d'augmenter la fidélité de la traduction du message génétique.
Notes et références |
(en) John G. Arnez et Thomas A. Steitz, « Crystal Structures of Three Misacylating Mutants of Escherichia coli Glutaminyl-tRNA Synthetase Complexed with tRNAGln and ATP », Biochemistry, vol. 35, no 47, 26 novembre 1996, p. 14725-14733 (PMID 8942633, DOI 10.1021/bi961532o, lire en ligne)
(en) P. Berg et E.J. Offengand, « An Enzymatic Mechanism for Linking Amino Acids to RNA », Proc. Natl. Acad. Sci. USA, vol. 44, 1958, p. 78-86 (PMID 16590176)
(en) Daniel Kern, André Dietrich, Franco Fasiolo, Michel Renaud, Richard Giegé et Jean-Pierre Ebel, « The yeast aminoacyl-tRNA synthetases: Methodology for their complete or partial purification and comparison of their relative activities under various extraction conditions », Biochimie, vol. 59, nos 5-6, juillet 1977, p. 453-462 (PMID 329894, DOI 10.1016/S0300-9084(77)80050-3, lire en ligne)
(en) Kelly Sheppard, Jing Yuan, Michael J. Hohn, Brian Jester, Kevin M. Devine et Dieter Söll, « From one amino acid to another: tRNA-dependent amino acid biosynthesis », Nucleic Acids Research, vol. 36, no 6, avril 2008, p. 1813-1825 (PMID 18252769, PMCID 2330236, DOI 10.1093/nar/gkn015, lire en ligne)
(en) Richard Giégé, « The early history of tRNA recognition by aminoacyl-tRNA synthetases », J. Biosci., vol. 31, 2006, p. 477-488 (PMID 17206068)
(en) Patrick O'Donoghue et Zaida Luthey-Schulten, « On the Evolution of Structure in Aminoacyl-tRNA Synthetases », Microbiology and Molecular Biology Reviews, vol. 67, no 4, décembre 2003, p. 550-573 (PMID 14665676, PMCID 309052, DOI 10.1128/MMBR.67.4.550-573.2003, lire en ligne)
(en) Michael Ibba et Dieter Söll, « Aminoacyl-tRNA Synthesis », Annual Review of Biochemistry, vol. 69, juillet 2000, p. 617-650 (PMID 10966471, DOI 10.1146/annurev.biochem.69.1.617, lire en ligne)
(en) Yuri I. Wolf, L. Aravind, Nick V. Grishin et Eugene V. Koonin, « Evolution of Aminoacyl-tRNA Synthetases—Analysis of Unique Domain Architectures and Phylogenetic Trees Reveals a Complex History of Horizontal Gene Transfer Events », Genome Research, vol. 8, no 9, août 1999, p. 689-710 (PMID 10447505, DOI 10.1101/gr.9.8.689, lire en ligne)
(en) M. Ibba et D. Söll, « Aminoacyl-tRNA synthesis », Annu. Rev. Biochem., vol. 69, 2000, p. 617-650 (PMID 10966471)
(en) Jiqiang Ling, Noah Reynolds et Michael Ibba, « Aminoacyl-tRNA Synthesis and Translational Quality Control », Annual Review of Microbiology, vol. 63, octobre 2009, p. 61-78 (PMID 19379069, DOI 10.1146/annurev.micro.091208.073210, lire en ligne)
Voir aussi |
Articles connexes |
- ARNt
- Acide aminé
- Ribosome
- Code génétique
 Portail de la biochimie
Portail de la biochimie


