Neurulation
Cet article est une ébauche concernant la biologie.
Vous pouvez partager vos connaissances en l’améliorant (comment ?) selon les recommandations des projets correspondants.
Consultez la liste des tâches à accomplir en page de discussion.
La neurulation est une étape du développement embryonnaire des métazoaires triploblastiques au cours de laquelle se met en place le système nerveux central. Elle succède à la gastrulation.
La neurulation se déroule en 2 étapes : la neurulation primaire et la neurulation secondaire.
La neurulation primaire comporte :
- la neuralisation : apparition de la plaque neurale par façonnage du neuroépithélium
- courbure et invagination
- fusion des bourrelets et fermeture du tube neural
- migration
La neurulation secondaire est « caudale ».
Chez les chordés, le système nerveux central est dorsal par rapport au tube digestif et à la chorde. La région dorsale de l'ectoderme s'épaissit pour former la plaque neurale et se referme en gouttière pour former un tube à l'origine de l'encéphale et de la moelle épinière. Les cellules à la frontière entre la plaque neurale et le reste de l'ectoderme donnent naissance aux cellules de la crête neurale qui sont à l'origine du système nerveux périphérique (et des mélanocytes).
Chez les invertébrés à l'exception des échinodermes, le système nerveux central est ventral par rapport au tube digestif (sauf dans la tête). Il est formé d'une série de ganglions reliés par des connectifs nerveux. La neurulation a surtout été étudiée chez la Drosophile. Des groupes de cellules isolées se détachent de l'ectoderme et donnent naissance aux ganglions.
Sommaire
1 Historique
2 Définition
2.1 Œufs télolécithes
2.2 Œufs hétérolécithes
3 L'induction de la neurulation
4 Articles connexes
5 Lien externe
Historique |

Matthias Schleiden
1838 -1839 : le botaniste allemand Matthias Schleiden et le physiologiste Theodor Schwann reconnaissent que tous les êtres vivants sont constitués de cellules, unité de base de la vie formée uniquement à partir de division d’autres cellules.
1924 : Hans Spemann et Hilde Mangold, embryologistes allemands, mettent en évidence l’induction chordo-mésodermique[Quoi ?].
1955 : Townes et Holtfreter démontrent que si on dissocie les cellules d’un embryon d’amphibien, il se forme des agrégats qui, selon la cellule de départ, forment un tissu ectodermique compact, mésodermique ou ectodermique.
Définition |
La neurulation correspond à la formation de ce qui sera plus tard le système nerveux chez les Métazoaires triploblastiques (ou triblastiques). Elle constitue l'étape qui précède l'organogenèse et qui suit ce que l'on appelle la gastrulation, au cours de l'embryogenèse.
Elle se déroule généralement sur la face dorsale de l'embryon dans le neuroectoderme, formé au cours de la gastrulation. La neurulation comporte plusieurs changements embryo-morphologiques par modification du neuroectoderme, notamment par la formation d'une plaque neurale et de bourrelets neuraux qui formeront la gouttière neurale. Par la suite, la fusion des replis neuraux (anciens bourrelets) formera le tube neural.
Œufs télolécithes |
Les œufs télolécithes (ou megalécithes) sont très gros, chargés en vitellus tassé vers le pôle végétatif et dont la quantité est importante par rapport à celle du cytoplasme formatif, de sorte que celui-ci, renfermant le noyau, se limite à un disque situé au pôle animal. Ils sont caractéristiques des oiseaux, des reptiles, et des céphalopodes. Leur neurulation est très proche de celle des œufs hétérolécithes, mais présente tout de même quelques particularités : leur rein vient de la région caudale du mésenchyme intermédiaire qui ne s’est pas divisé et a ainsi créé un blastème.
Ils ne possèdent pas d'archentéron, mais une gouttière renversée est formée par soulèvement de la lame entoblastique intra-embryonnaire. Celle-ci n’est pas entièrement refermée, mais après apparition des plis neuraux, elle se refermera progressivement vers l’avant pour s’achever au niveau caudal.
Œufs hétérolécithes |
Les œufs hétérolécithes sont assez gros, pauvre en vitellus avec un net gradient vitellin hétérogène (la concentration en vitellus est plus important au pôle végétatif) et anisotrope (tous les axes de l’œuf ne sont pas identiques). Ils sont caractéristiques des amphibiens, des annélides, et des lamellibranches.

Schéma d'une fin de neurulation sur un œuf hétérolécithe.
La neurulation des œufs hétérolécithes comporte 4 phases :
- Epaississement de l'ectoderme dans la région médiane et formation de la plaque neurale.
Pendant que la fente blastoporale est en train de se former, l’ectoderme qui se situe sur la partie médiane de l’embryon va subir un aplatissement ainsi qu’un épaississement. Celui-ci se propagera latéralement et progressera sur toute la longueur de la phase dorsale de l’embryon. Grâce à ce mécanisme la plaque neurale sera mise en place.
La région antérieure, alors élargie, constituera la plaque neurale dite céphalique, et se prolongera en une partie qui sera quant à elle plus étroite : c'est la plaque neurale médullaire.
À leur tour, les bords latéraux de la plaque neurale s’épaissiront et se soulèveront selon l’axe dorso-ventral de la neurula (embryon en début de phase de la morphogenèse). Ils formeront les bourrelets neuraux céphaliques ou médullaires selon leur position.
- Soulèvement des bourrelets et soudure de la plaque et formation de la gouttière neurale.
Un soulèvement des bourrelets ainsi qu’une courbure de la plaque neurale se produira, ce qui conduira à la formation de la gouttière neurale, qui formera le tube neural.
- Fusion des replis neuraux.
Les bourrelets neuraux vont fusionner (impliquant l’expression des molécules d’adhésion ou de fusion) et se propager vers les régions antérieures et postérieures de la neurula. Le sillon neural sera ainsi formé dans la région dorsale.
- Fermeture du tube neural et formation du cerveau.
Le tube neural, situé sous l’ectoderme et constituant le système nerveux central primitif, va progressivement se différencier selon l’axe antéro-postérieur. Il formera trois vésicules du cerveau initial : le cerveau antérieur (prosencéphale), moyen (mésencéphale) et postérieur (rhombencéphale).
Il sera aussi à l’origine de la moelle épinière : les deux extrémités de celui-ci, le neuropore antérieur et le neuropore extérieur, se rapprocheront au niveau de l’endoderme, ce qui contribuera à former la voûte de l’archentéron. Les lames latérales se rapprocheront dans la région ventrale et formeront le cœur dans la région antérieure.

Début de la neurulation.
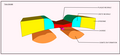
Mouvements convergents qui poussent à la formation des crêtes neurales. Apparitions de somites.


L'induction de la neurulation |
- Expérience de Spemann et Mangold
Spemann et Mangold, qui étaient deux embryologistes allemands, ont révélé en 1924 par leur expérience le rôle inducteur des cellules présentes au-dessus de la lèvre dorsale du blastopore de Xénope, qui est équivalent au nœud de Hensen chez les vertébrés. Ces cellules sont appelées aujourd’hui « Centre Organisateur de Spemann ». Ils ont prélevé des cellules d'une jeune gastrula puis les ont greffées sur d'une autre gastrula d'une espèce proche mais de pigmentation différente, au même stade de développement, sur sa partie ventrale. Le greffon s’invagine et s’incorpore dans le zygote, puis, lors de la neurulation, une plaque neurale secondaire en position ventrale apparaît en plus de la plaque neurale primaire apparue en position dorsale normalement au cours du développement, d'où le terme de « double neurulation ». Les deux plaques neurales vont alors évoluer indépendamment et constituer chacun un tube neural au terme d'un mécanisme complexe expliqué ci-dessous.
Cette expérience a conduit à la formation d'un double têtard, dont l'un est issu du greffon qui possède des pouvoirs neurogène et mésoblastogène. Son pouvoir neurogène se traduit par capacité du greffon à induire la formation d'un système nerveux complet et d'un encéphale qui lui sont propres à partir des cellules de l'ectoderme provenant de l'embryon ayant reçu le greffon. Quant à son pouvoir mésoblastogène, il permet aux cellules du greffon de recruter des matériels cellulaires du receveur, qui vont se différencier en matériels mésodermiques afin de compléter la formation de l'embryon à partir du greffon. Le greffon est également capable d'auto-différenciation puisqu'en conditions normales, les cellules de la lèvre dorsale du blastopore dont il est issu donnent une partie de l'endoderme pharyngien et quelques éléments de la corde et des somites. Cette expérience nous montre le rôle inducteur du tissu présent sur la partie dorsale du zygote ainsi que son caractère déterminé, et plus particulièrement les cellules situés au-dessus de la lèvre dorsale du blastopore : le Centre Organisateur de Spemann.
- Expérience de Saxen et Toïvonen
Souvent, plusieurs sources d'inducteurs sont présentes simultanément ou successivement pour provoquer une induction . Sulo Toïvonen et Lauri Saxen ont montré en 1955 que, lors de l'induction neurale, la formation du système nerveux axial nécessite l'action coordonnée de deux inducteurs : ils implantent simultanément dans le blastocoele d'une jeune gastrula un morceau de foie de cobaye et de la moelle osseuse. Le blastula se développe alors normalement, mais il se forme aussi du cerveau moyen et du cerveau postérieur ainsi que de la moelle épinière par induction de la part des implants. De plus, ils remarquent que plus l'implant est conséquent, plus le système nerveux sera développé.
- Expérience de Saxen et Coll
Ce modèle est précisé par l'expérience de Saxen et Coll en 1964, qui associent l'ectoderme avec chacun de ces 2 inducteurs, séparément puis ensemble.
Après 24 heures d'induction, les cellules ectodermiques dissociées se ré-agrègent pour former respectivement le cerveau postérieur en présence du morceau de foie de cobaye et la moelle osseuse en présence de la moelle épinière, mais en présence des 2 en même temps, elles forment aussi le cerveau moyen (plus tard) en plus du cerveau postérieur et de la moelle épinière.
L'induction se fait donc séparément pour former la moelle épinière et le cerveau supérieur, et simultanément pour former le cerveau moyen.
L'induction se fait donc en 2 étapes :
- Induction neurale céphalique et postérieure
Les inductions émises par le centre organisateur de Spemann et Mangold pendant la gastrulation (par invagination sous l'ectoderme) forment la plaque neurale, puis le tube neural, sous l'action de 2 signaux:
- Le premier signal, émis dans l'ectoderme dorsal par le mésoderme grâce à des protéines, induit la différenciation de structures antérieures de type prosencéphale.
- Le second signal, émis lui aussi dans l'ectoderme dorsal par la lèvre blastoporale, permet la formation de structures postérieures le long de l'axe céphalo-caudal (tube neural).
- Induction de l'épiderme et différenciation neurale
L'action des protéines provoquant la différenciation des cellules ectodermiques en épiderme est inhibée par les inducteurs neuraux: c'est la différenciation neurale par défaut.
D'après les expériences de Spemann et Mangold, l'ectoderme forme du système nerveux quand ils sont actifs et forme de l'épiderme quand les inducteurs neuralisants sont inactifs, c'est-à-dire à la fin de la gastrulation.
Articles connexes |
- embryogenèse
- gastrulation
Lien externe |
- « La Neurulation »
 Portail de la biologie
Portail de la biologie  Portail des neurosciences
Portail des neurosciences


